
1. INTRODUCTION
On a global scale, there is compelling evidence that the temperature of the Earth is rising, the atmosphere is drying up, and extreme weather events are intensifying as a consequence of climate change induced by anthropogenic factors. In Korea, rising temperatures and precipitation have altered the behavior of species within local ecosystems. Furthermore, these impacts are influencing the population dynamics of termites in Korea. The prevalence of plant-decomposing termites has increased significantly across the country, with the active period of dominant termites in Korea exhibiting a notable 16-day increase (Kim et al., 2023a). As a result, the incidence of damage to wooden structures, particularly those of cultural heritage value, is on the rise. The termite species inhabiting Korea have been recorded to include Reticulitermes speratus kyushuensis, Reticulitermes speratus speratus, and Reticulitermes kanmonensis. In addition, Glyptotermes nakajimai was discovered in Yeoseodo in 2021. In 2023, the presence of Cryptotermes domesticus was confirmed within a residential structure in Gangnam, Seoul, while Incisitermes minor was identified in Changwon, Gyeongnam (Kim, 2023). Kim et al. (2023b) used species distribution modeling techniques for C. domesticus to predict overall low climate suitability for Korea but reported its potential for establishment and spread.
The “Surveys on Species Affecting Wooden Cultural Properties” in 2016 to 2019 confirmed termite detection dog reactions in 87.6% of cultural properties, with visible termite damage in 51.1%. In particular, wooden cultural heritage in Jeollabuk-do and Jeollanam-do had a higher degree of termite damage than wooden cultural heritage in the Seoul metropolitan area, Chungcheongbuk-do, and Gyeongsangbuk-do (Kim and Chung, 2022). Bak et al. (2023) conducted an analysis of the cases and problems associated with termite damage to traditional buildings with the objective of preserving the distinctive characteristics of hanoks while developing a construction method that is compatible with modern requirements. In light of the critical need to safeguard wooden cultural heritage from termite infestation, a range of techniques is being employed, including X-ray inspections (Lee and Kim, 2022).
The infestation of wooden structures by termites is not a phenomenon exclusive to Korea; it is also observed in other countries around the world. It has been confirmed that the Forbidden City in China is infested with termites. In order to control the infestation, chemical controls have been implemented for the species found (Zhang et al., 2022). Additionally, instances of termite infestation have been documented in conventional wooden structures across the globe. In Australia, the incidence of termite damage to wooden houses is rising annually. In Indonesia, termite attacks have resulted in economic losses amounting to IDR 8.7 trillion per year (Subekti et al., 2024). Accordingly, there is ongoing research into the distribution and behavior of wood-damaging termite species (Reid, 2009). Even Japan, where about 58.9% of homes are made of wood, suffers from both wet and drywood termites (Seo, 2016). In Japan, soil and wood members are treated with eco-friendly pesticides, and research is being conducted into termite ecology with a view to implementing an appropriate control strategy (Tsunoda, 2005). Based on the expectation that global trade and global warming would facilitate the movement of termites, Duquesne and Fournier (2024) modeled the invasion distributions of 10 termite species, reporting that termites were projected to increase in the tropics and subtropics, as well as in temperate regions.
Subterranean termites (Reticulitermes spp.) live in damp, low-light conditions and go underground when temperatures drop (Han et al., 1998). The prevention of subterranean termite damage requires the identification of termite colonies and the extent of their activity (Lee et al., 2023). There are five known species of termites in Korea. Nevertheless, no study has yet examined and analyzed the distribution of subterranean termites by region across South Korea. Therefore, this study aims to analyze termite species living in 85 regions of Korea.
There is a pervasive initiative across both the public and private sectors to advance wood construction as a means of curbing carbon emissions and achieving the national goal of net-zero emissions by 2050. The survey on the distribution of termites on the Korean Peninsula in this study is expected to provide the basic data for preventing termite damage to wooden structures in Korea.
2. MATERIALS and METHODS
In order to investigate the distribution of termites in Korea, termite samples were collected from the regions where the Korea Meteorological Administration (KMA) collects climate data. The termites collected from 85 regions in Korea, including Cheorwon, the northernmost province, and Jeju Island, the southernmost province, were analyzed for their species (Table 1). Termites were collected from a forest or forest-adjacent site in each region, with three colonies per site. The goal was to collect all worker, soldier, and queen termites from a single colony. Damaged trees were collected together, recording the address, elevation, latitude, and longitude of the collection site. Sampling was conducted from late September to early November 2023, a period of relatively low termite activity, which may have contributed to the difficulty of sampling, but it is unlikely to have contributed to differences in species abundance.
All termites collected for species identification were stored in 99% EtOH to prevent damage to their DNA.
For analyzing the morphological characteristics, dried specimens were prepared from soldier type termites, photographed with a Leica Microscope (MSV266, Leica, Wetzlar, Germany), and images were synthesized with the Zerene stacker (Zerene System, Richland, WA, USA). The length and width of the head as well as the length of the mandibles were measured, and morphological characteristics of the head, mandibles, antennae, pronotum, and forelegs were observed.
For genetic analysis of termites, the middle and hind legs on one side of the termites were crushed with a crushing rod. The DNA extraction process followed the protocol of the DNeasy® Blood & Tissue Kit from Quiazen (San Diego, CA, USA). PCR was performed using primers such as COI; LCO1490, HCO2198 (Folmer et al., 1994), COII; TL2-J-3037 (Liu and Beckenbach, 1992), and TK-N-3785 (Simon et al., 1994), as shown in the following table. Subsequently, the species was identified through DNA sequencing, with the resulting sequence then compared to those recorded in GenBank (Table 2). In order to analyze the genetic differences between the populations, a phylogenetic tree analysis was performed with the MEGA11 program using the mitochondrial genes (COI: 658 bp and COII: 747 bp, 742 bp) used for species identification. Phylogenetic tree analysis was performed by maximum likelihood using the GTR + G + I model (bootstrap rep. 1000).
| Primer name | Primer sequence (5’→3’) | Sense |
|---|---|---|
| LCO1490 | GGTCAACAAATCATAAAGATATTGG | Forward |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | Reverse |
| TL2-J | ATGGCAGATTAGTGCAATGG | Forward |
| TK-N | GTTTAAGAGACCAGTACTTG | Reverse |
3. RESULTS and DISCUSSION
There were four species of termites found in the 85 regions examined in this study: R. speratus kyushuensis, R. kanmonensis, R. speratus speratus, and I. minor. The 216 termites classified were composed of 187 R. speratus kyushuensis, 27 R. speratus speratus, and 1 R. Kanmonensis, and 1 I. minor (Figs. 1 and 2). Although R. speratus kyushuensis, the Japanese termite, has been known to inhabit Korea for a considerable time, there are no records of its first appearance. R. kanmonensis was discovered in Jeollabuk-do and Chungcheongnam-do in 1999 and documented as morphologically and genetically distinct from R. speratus kyushuensis (Lee et al., 2015). R. speratus speratus is a subspecies discovered in 2023 and has been confirmed to have genetic differences from R. speratus kyushuensis, the dominant species in Korea (Lee et al., 2023). I. minor analyzed in this study, commonly referred to as the western drywood termite, is notable for its dietary preference for drywood, a trait that distinguishes it from other termite species analyzed in this study.


The northern and central regions of Korea are mostly inhabited by R. speratus kyushuensis. R. speratus speratus has been found to inhabit Ganghwa in the northern region and the southern regions of the country with relatively high temperatures. There were 11 regions where R. speratus kyushuensis and R. speratus speratus species were found together. The only R. kanmonensis found in this study inhabited Gunsan along with R. speratus kyushuensis and R. speratus speratus (Fig. 3). In 2023, damage caused by I. minor was confirmed in a facility (outdoor gazebo) in an apartment complex in Jinhae-gu, Changwon-si, Korea (Lee et al., 2024). This species has evolved a number of adaptations that enable it to thrive in arid environments, such as the ability to obtain moisture from trees and to produce water through oxidative metabolism. It is also characterized by producing pellets weighing 0.1 to 0.15 g (Cabera and Scheffrahn, 2001). Previous studies have shown that it selectively targets the sapwood first (Khoirul, 2017).
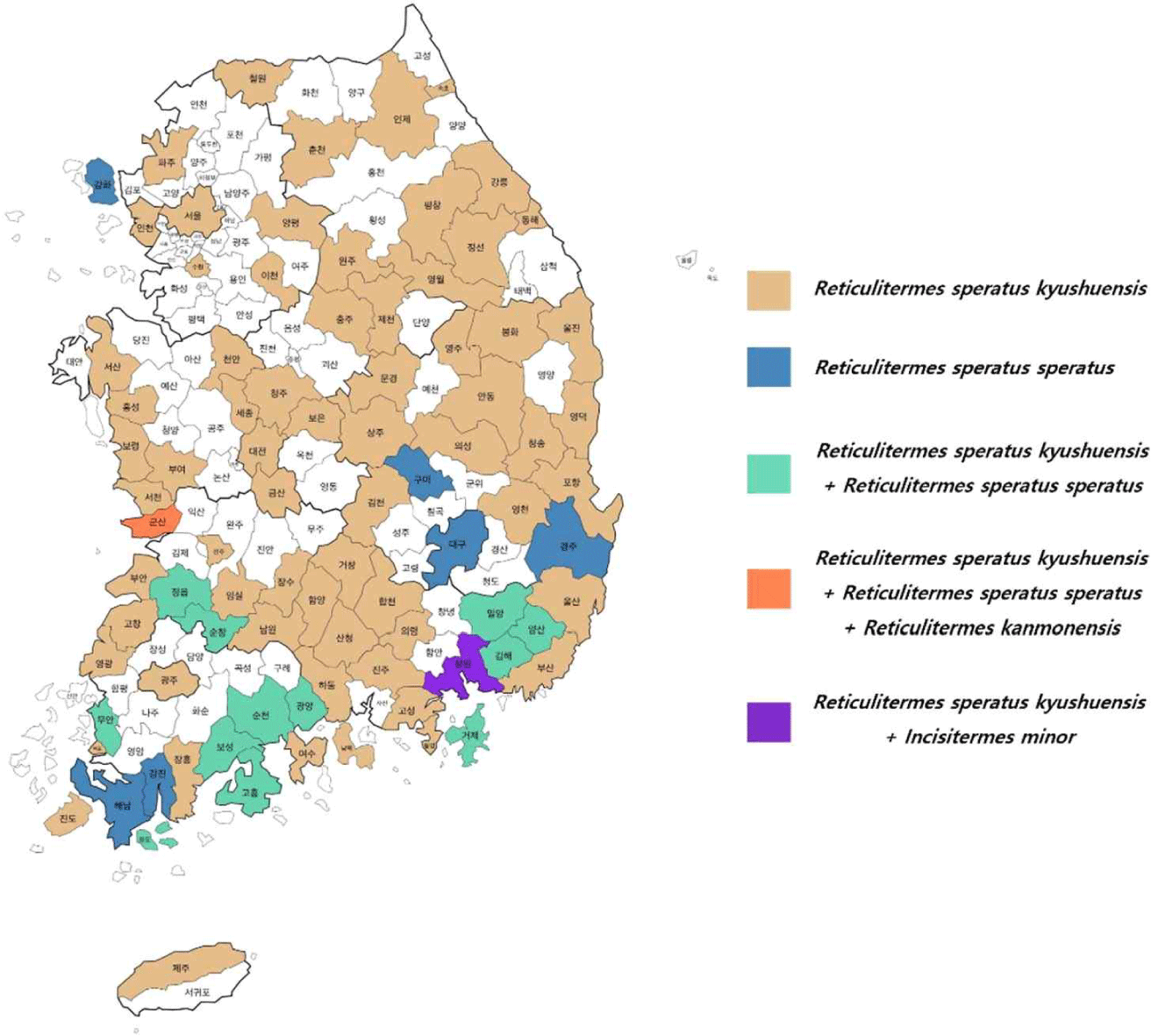
The lowest temperatures in winter and the highest temperatures in summer have a huge impact on termite colonization (Houseman et al., 2001). At the coldest temperatures, termites adjust their carbohydrate intake and metabolism to balance body water content and lipids for survival (Choi et al., 2016). The average monthly temperature (Fig. 4) from 1991 to 2020 in the regions where termites were collected in this study was the lowest at –6.9°C in January in Pyeongchang and the highest at 27.4°C in August in Yangsan. In terms of temperature, termite (R. speratus kyushuensis) survival was highest at 25°C, with no survival above 35°C and below –4°C (Lee and Jeong, 2004). I. minor thrives at temperatures around 35°C, and no temperature limits have been clearly identified (Indrayani et al., 2007). Since the average maximum temperature in all regions did not exceed 35°C, there were no regions where high temperatures prevented termites from inhabiting. The lowest temperature in January was lower than –4°C in Pyeongchang, Paju, Jecheon, Inje, Cheorwon, and Chuncheon, which would make it difficult for termites to survive in some regions (Fig. 4). A previous study examined the natural habitat of termites and revealed that they create subterranean habitats beneath tree roots to withstand external environmental conditions (Takata et al., 2023). Therefore, it is unlikely that the habitat of subterranean termites is affected by average temperatures, as they avoid high temperatures by creating underground ant trails during the hot summer months and live underground during the winter months.
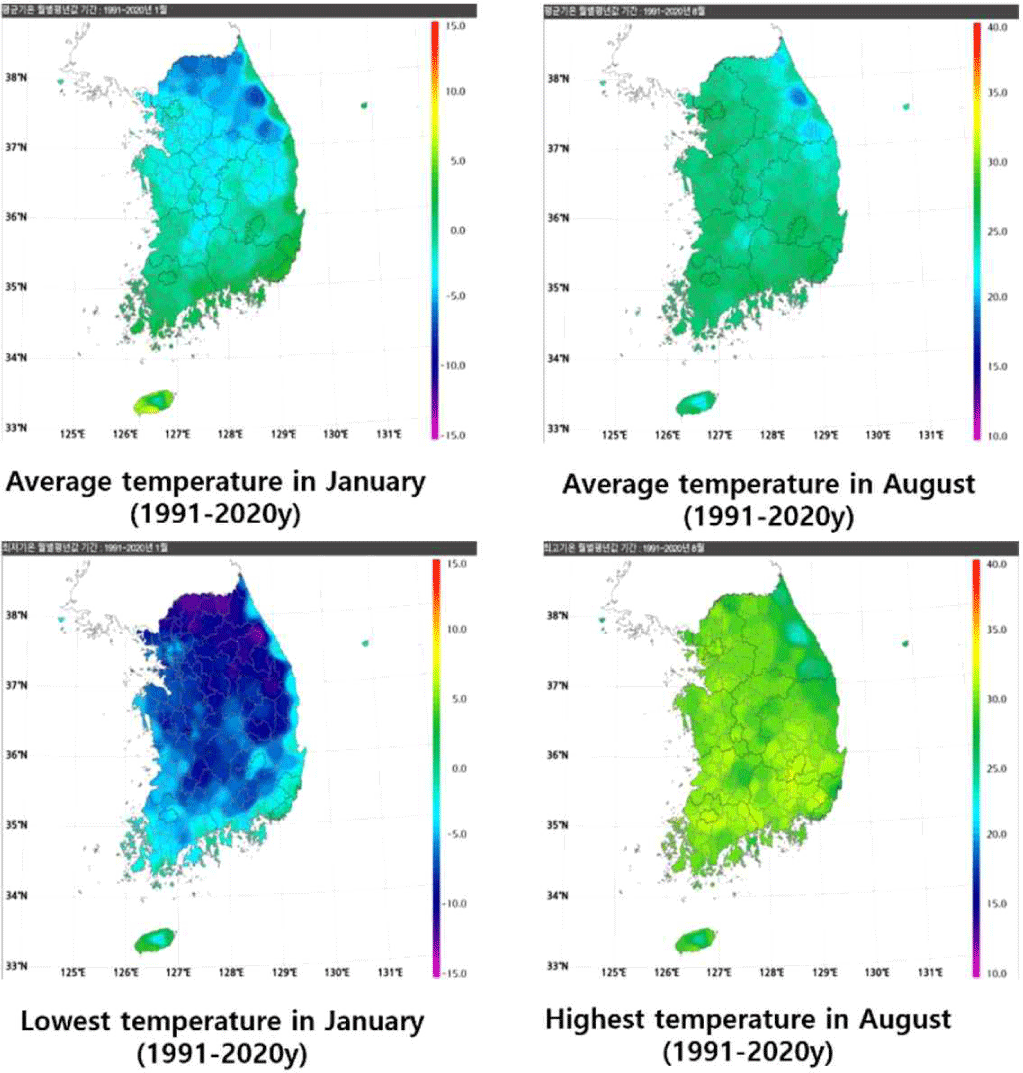
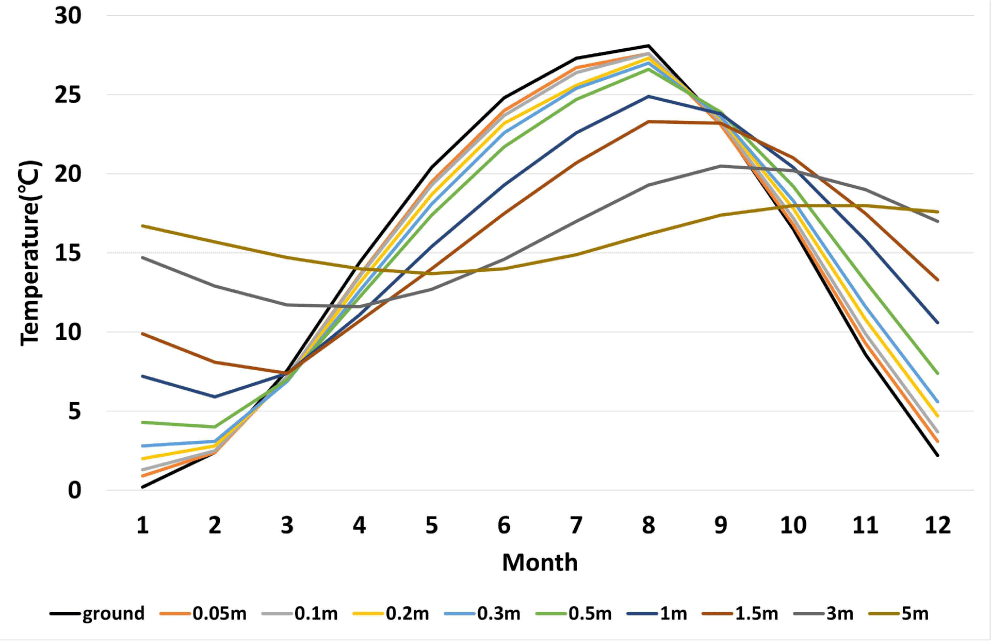
In Korea (survey regions; from ground surface to –0.3 m: Chuncheon, Gangneung, Seoul, Incheon, Suwon, Seosan, Daejeon, Busan, Yeosu, and Jeju; from –0.5 to –5 m: Chuncheon, Gangneung, Seoul, Incheon, Daejeon, Busan, Yeosu, and Jeju), the average monthly underground temperature from 1973 to 2023 was the lowest in January at 0.2°C. The highest temperature was recorded in August at 28.1°C. The temperature at 0.05 m below ground, close to the surface, was the lowest in January at 0.9°C and the highest in August at 27.6°C. The temperature at 5 m below ground, the deepest depth, had the smallest difference between low and high temperatures at 13.7°C–18.0°C. Therefore, the deeper termites go into the ground, the less likely they are to suffer from cold damage in the winter (Fig. 5). This is in line with a study that reported that termites of the genus Reticulitermes can avoid the cold in winter by living at a depth of more than 1 m below ground (Cabrera and Kamble, 2001).
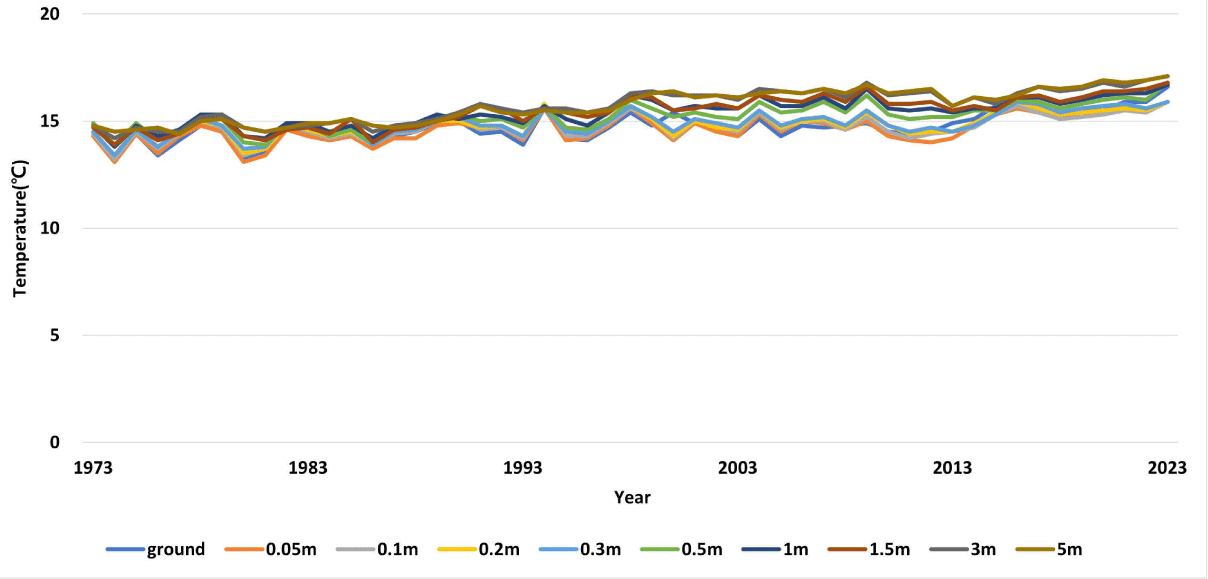
From 1973 to 2023, the average annual ground temperature tended to increase due to climate change, with temperatures at 5 m below ground recorded at 13.1°C–17.1°C (Fig. 6). The ground surface and ground depth did not show a clear relationship, with lower temperatures mostly at and near the ground surface and higher temperatures measured at 0.05 to 5 m below ground.
The presence of moisture is a significant contributing factor to the colonization of termites. Termites of the genus Reticulitermes determine their diet based on the temperature and moisture of the soil (Janowiecki and Vargo, 2021). The relative humidity of the atmosphere in Korea, which affects termite colonization, has been gradually decreasing. In other words, dryness is increasing as a result of climate change (Fig. 7).
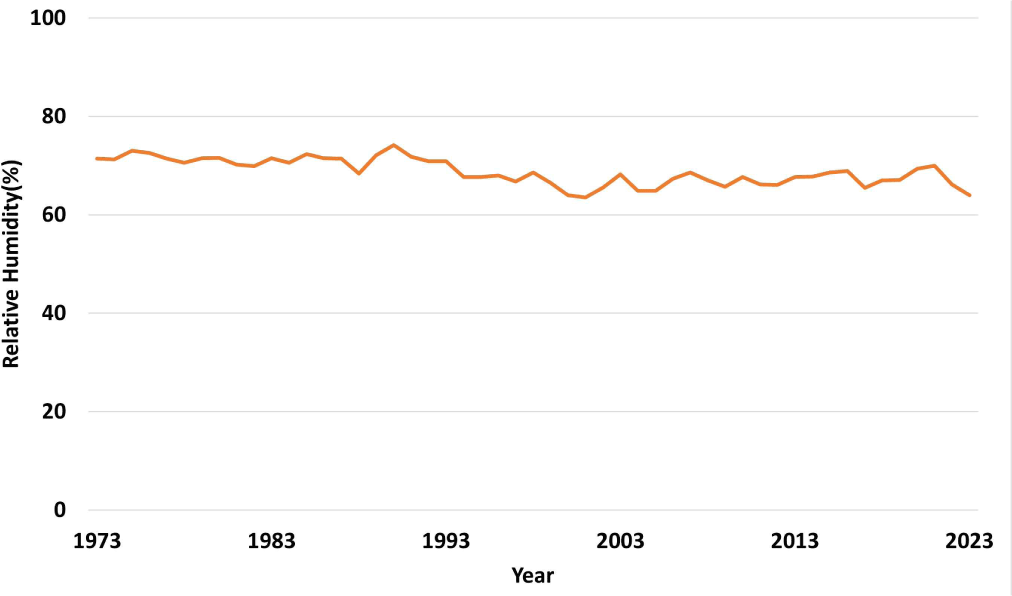
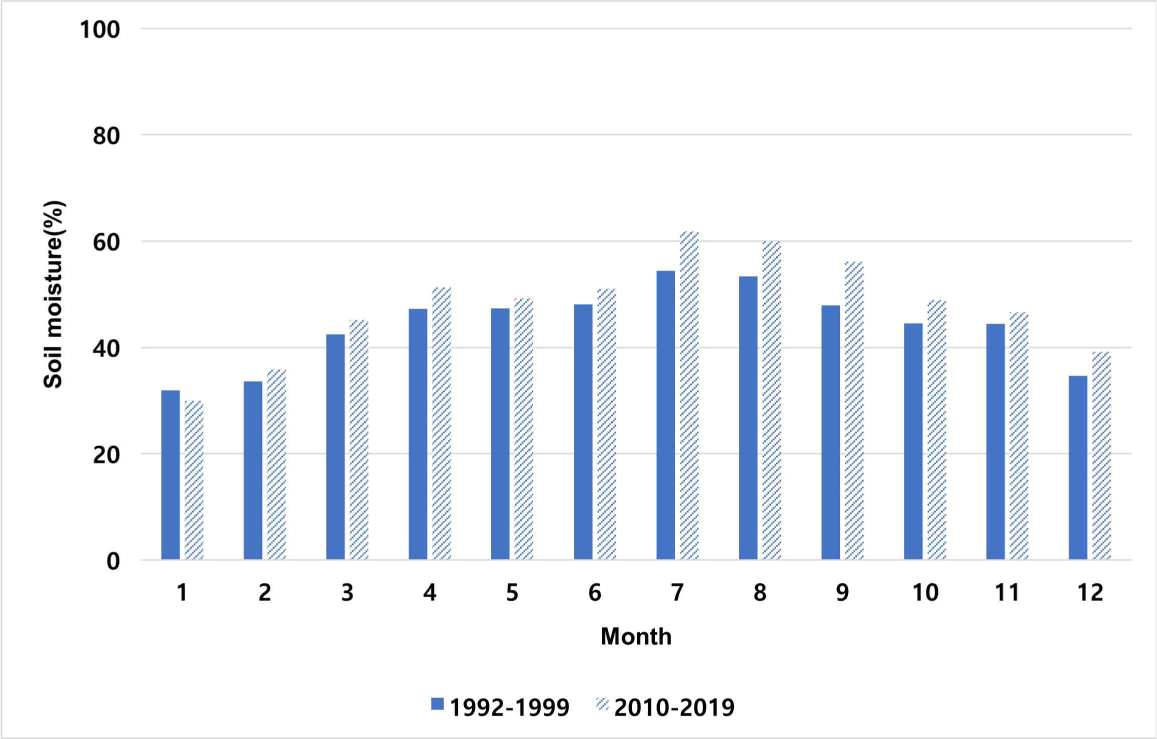
In the 1990s and 2010s in Korea, soil moisture was reported to be low in January and December and high in July and August. Soil moisture is increasing over time, with all months except January in the 1990s showing higher moisture in the 2010s compared to the 1990s (Fig. 8). Soil moisture causes ecosystem changes (Deng et al., 2020). Accordingly, fluctuations in soil moisture were postulated to exert an influence on the subterranean ecology of termites.
The Shared Socio-economic Pathways climate change scenarios reported by the KMA indicate that temperatures and precipitation in Korea will gradually increase under all scenarios. In light of the predicted alterations to the climate, it is anticipated that there will be changes in the species and numbers of termites in Korea, which means that the impact of termites on wood or wood structures is likely to intensify, exceeding its current magnitude.
The recent identification of subterranean termites and western drywood termites infesting drywood in Korea underscores the necessity for the formulation of control measures tailored to the distinctive characteristics of each termite species. Termites can be controlled using chemical or physical methods. Due to the temporary nature and human health risks of chemical control, physical methods like baiting and thermite mashing have been suggested (Gu and Cheon, 2018). For physical control of domestic termites, research is being conducted on the minimum passage diameter of termites to develop termite entry barriers (Kim et al., 2020). Furthermore, monitoring methods such as the use of dyes to identify termite territories are being used (Im and Han, 2021). In the United States, there are many different types of termites, including subterranean, drywood, and wetwood termites, requiring different control methods depending on the characteristics of the termite (Jeong, 2011). Recently, research has been conducted on the potential use of plant stem extracts as an environmentally friendly control method (Zalsabila et al., 2024). There are currently six species of termites identified in Korea: R. speratus kyushuensis, R. speratus speratus, R. kanmonensis, G. nakajimai, C. domesticus, and I. minor. As the species of termites that can live and the number of active days are expected to increase due to climate change, it is imperative to implement eco-friendly and effective control measures to prevent termite damage that could impede the revitalization of wooden structures in Korea.
4. CONCLUSIONS
This study examined the distribution and species of termites in 85 locations across the country in an effort to prevent termite damage in Korea. The presence of termites was confirmed in all of the surveyed areas, with four species identified: R. speratus kyushuensis, R. speratus speratus, R. kanmonensis, and I. minor. These species were observed to inhabit the sites either individually or in combination, rather than as a single species. Even in winter, when Korea records the lowest temperatures, termites are still able to live in the winter as the temperature at 1 m below ground level is conducive to the survival of termites. As temperatures in Korea rise, the number of active days and terrestrial activities of termites is likely to increase, accompanied by a corresponding rise in the damage they cause. It is anticipated that the effects of climate change and increased international trade will result in an increase in the activity of existing termites as well as the emergence of new species. It is, therefore, imperative that a comprehensive plan be implemented to control the spread of termites, which present a significant risk to the stability of wooden structures.